
Ontogeneze desetinohých korýšů


Ontogeneze, tedy vývoj jedince od oplození vajíčka do dospělosti se u desetinohých korýšů liší podle prostředí, na které se adaptovali, a tedy podle délky larválního vývoje. Většina desetinohých korýšů má nepřímý vývoj s proměnou nedokonalou, což znamená, že z larvy se stává dospělec bez stádia kukly a bez metamorfózy (mylná domněnka, že korýši prodělávají metamorfózu, byla definitivně vyvrácena již v roce 1840).
Po každém svléknutí krunýře larva postoupí do dalšího vývojového stadia. Na rozdíl od hmyzu s tímto typem proměny však larvy korýšů nejsou zpočátku podobné dospělci, naopak se od něj morfologicky velice liší. Proto nejsou zdaleka všechny larvy nacházené v zooplanktonu v mořích přiřazeny ke konkrétním druhům desetinohých korýšů. Přiřazení larev k dospělcům komplikuje odlišné prostředí, ve kterém obě stadia některých druhů žijí (moře vs. sladká voda či terestrické prostředí) a zatím nepopsané bentické hlubokomořské druhy, jejichž dospělci oproti planktonním larvám často unikají pozornosti. Například larvy listorožce velkého (Scyllarides latus) byly identifikovány až v roce 2013, ačkoliv druh jako takový byl vědecky popsán již v roce 1803.
První kompletní ontogenetický vývoj u desetinohého korýše byl popsán v roce 1870 u humra amerického (Homarus americanus) a jeho autorem byl americký zoolog Sidney Irving Smith. Problematické je zařazení druhů s potlačeným larválním vývojem (raci, někteří krabi, některé krevety), jejichž larva prodělává většinu či celý vývoj uvnitř vajíčka. Vzhledem k tomu, že tyto druhy nemají volně pohyblivou planktonní larvu, jsou většinou řazeny mezi druhy s vývojem přímým.
U druhů s dlouhým (standardním) larválním vývojem se z vajíčka líhne drobná planktonní larva (o velikosti přibližně 2 mm), která je volně pohyblivá a nazývá se nauplius. Z ní se pak stává metanauplius, protozoea, mysis a tzv. post-larva neboli mastigopus (u garnátů). Zvláštností u larvy garnátů ve stadiu mysis je přítomnost párových statocyst na ocasní ploutvičce. Stadium mysis se u garnátům příbuzných korýšů z čeledi Sergestidae nazývá acanthosoma (tedy stejně jako jeden rod knězovitých ploštic). Larva se může líhnout již jako tzv. zoea (u krabů, krevet a poustevníčků) či phyllosoma (u langust a listorožců), což odpovídá zmíněnému stadiu mysis u garnátů. Larvy ve stadiu nauplius a metanauplius mají jen jedno tzv. naupliové oko, které detekuje pouze směr a intenzitu světla. Postupně toto oko zaniká a tvoří se párové složené oči. Planktonní larvy se pohybují pomocí antén a brv. Abdomen mohou mít zakončen různými útvary (např. vidlicí u krabů nebo ocasní ploutvičkou u poustevníčků a garnátů). Pro koordinované plavání určitým směrem je larva na krunýři většinou vybavena různě dlouhým rostrálním a posteriorním trnem. Po poškození či ablaci těchto trnů plave larva chaoticky a její schopnost manévrovat a couvat je tím velice omezená.
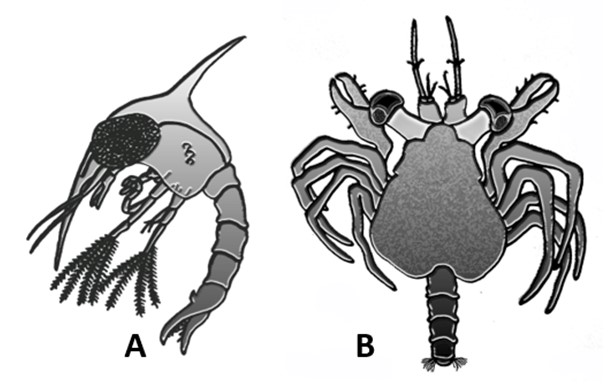
A) planktonní larva kraba zvaná zoea není dospělci podobná; B) bentická larva megalopa se dospělci podobá více než zoea, ale má stále ještě napřímený abdomen a v poměru k tělu velké oči
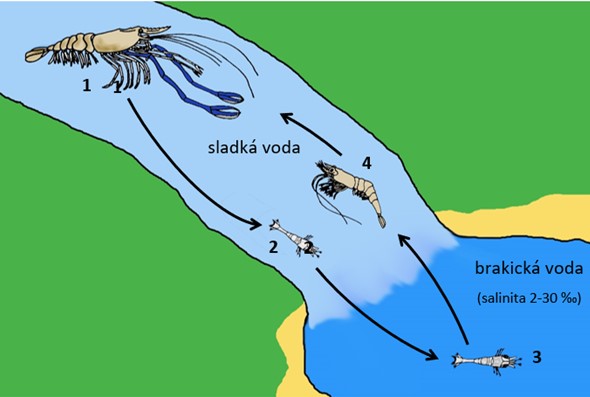
Životní cyklus krevetek Rosenbergových (Macrobrachium rosenbergii): 1) dospělí jedinci jako zobrazený samec žijí v tekoucí sladké vodě, samice po nakladení nosí vajíčka přitmelená k pleopodům (20-21 dní); 2) vylíhlá planktonní larva zoea driftuje s proudem do moře (do 48 hodin); 3) larvální vývoj v moři (11 stadií, 15-40 dní); 4) bentická post-larva zvaná parva migruje zpět do sladké vody a mění se na juvenilní krevetku, která dospívá za 120-210 dní. Velikostní poměr vývojových stadií na obrázku neodpovídá skutečnosti. V závorce je u každého vývojového stadia uvedena doba, po kterou korýš v tomto stadiu setrvává
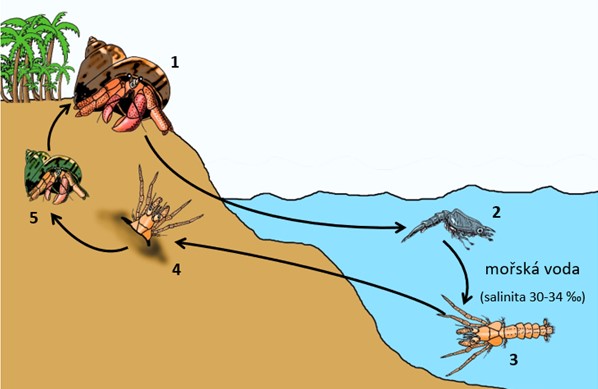
Vývojový cyklus suchozemských poustevníčků rodu Coenobita: 1) gravidní samice nosí vajíčka na pleopodech (25-29 dní); 2) samice vajíčka těsně před líhnutím vypouští do moře, kde při kontaktu se slanou vodou praskají a uvolňují se z nich planktonní larvy zoea (2-7 stadií, 17-80 dní); 3) bentická larva glaucothoe (21-28 dní); 4) glaucothoe vyhledá prázdnou ulitu plže, ve které ukryje zadeček, migruje na souš, zahrabe se a mění se na juvenilního poustevníčka (3-4 týdny); 5) juvenilní poustevníčci opouštějí nory a dospívají za další dva roky. Velikostní poměr vývojových stadií na obrázku neodpovídá skutečnosti. V závorce je u každého vývojového stadia uvedena doba, po kterou korýš v tomto stadiu setrvává
Planktonní larva prochází jedním i více stadii a mění se na přibližně 5 mm velkou post-larvu zvanou parva či mastigopus (u krevet a garnátů), megalopa (u krabů), glaucothoe (u poustevníčků) či puerulus též označovaná jako nisto nebo pseudibacus (u langust a listorožců). Megalopa krabů ještě nemá zadeček stočený pod hlavohruď jako je tomu u juvenilních a dospělých jedinců. V tomto stadiu mizí naupliové oči a objevuje se dvojice složených očí. Larva, která prochází opět jedním či několika stadii, má již stejný počet končetin jako dospělec a morfologicky je mu podobná mnohem více než larva planktonní. Tato larva může také plavat, ale převážně se pohybuje po dně, je tedy bentická. U humrů se jednotlivá vývojová stadia označují pouze číslem I-IV. První tři stadia jsou pelagická (larvy plavou ve vodním sloupci), čtvrté je bentické.
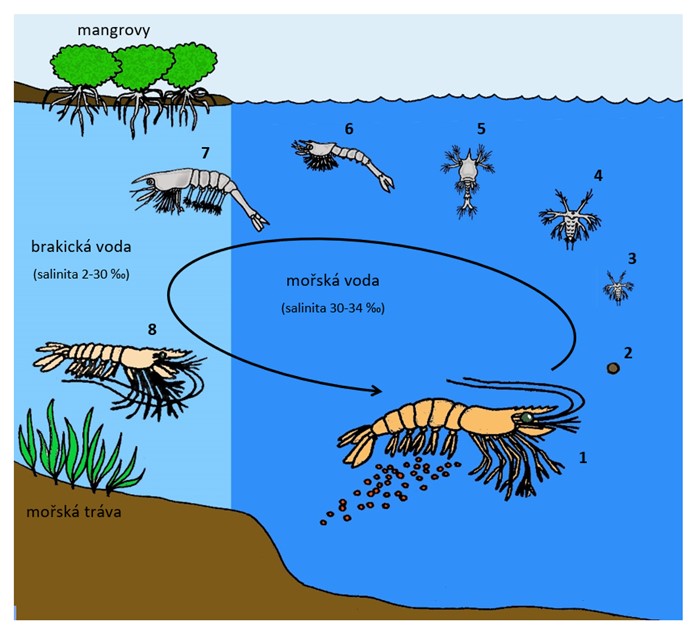
Životní cyklus pelagických garnátů rodu Penaeus (podřád Dendrobranchiata): 1) dospělci žijí v moři, samice vypouští vajíčka přímo do vody; 2) vajíčko (12-24 hod); 3) nauplius (3 stadia, 1 den); 4) metanauplius (2-3 stadia, 1 den); 5) protozoea (3 stadia, 3-5 dní); 6) mysis (3 stadia, 3-5 dní); 7) post-larva (mastigopus) migruje do brakické vody (3-35 dní); 8) juvenilní jedinec ukrývající se v porostech mořské trávy (2,5-10 měsíců), pak migruje do moře a stává se z něj dospělec. Velikostní poměr vývojových stadií na obrázku neodpovídá skutečnosti. V závorce je u každého vývojového stadia uvedena doba, po kterou korýš v tomto stadiu setrvává

Životní cyklus mořských krabů (infrařád Brachyura): 1) gravidní samice se snůškou vajíček na pleopodech (6-25 dní); 2) planktonní larva zoea (4-7 stadií, 12-24 dní); 3) bentická post-larva megalopa (1 stadium, 5-7 dní); 4) juvenilní krab (4-18 měsíců). Velikostní poměr vývojových stadií na obrázku neodpovídá skutečnosti. V závorce je u každého vývojového stadia uvedena doba, po kterou korýš v tomto stadiu setrvává
Většina larválních stadií desetinohých korýšů byla původně popsána jako samostatné druhy a rody. Kupříkladu v roce 1785 Otto Friedrich Müller zařadil planktonní larvy do dnes již neplatného rodu Nauplius. V roce 1802 Louis Augustin Guillaume Bosc popsal larvy krabů jako dnes neplatný samostatný rod Zoea. Oba názvy tedy přetrvaly jen v podobě označení příslušných larválních stadií. U tropických druhů trvá larvální vývoj většinou méně než dva týdny, zatímco u druhů z chladnějších oblastí může trvat i déle než měsíc. Rekordmany jsou langusty a listorožci, jejichž larvy a post-larvy setrvávají v těchto stadiích až dva roky. Planktonní larvy se živí zoo- a fytoplanktonem a vlivem proudění mořské vody se mohou šířit po velkém areálu daleko od rodičů a mohou zakládat nové populace, případně obnovovat populace zdecimované. Například larvy krabů a poustevníčků byly dokonce objeveny u břehů Antarkitidy, konkrétně u ostrova Krále Jiřího (největší ostrov ze souostroví Jižní Shetlandy). Oproti tomu mláďata druhů s potlačeným larválním vývojem nemohou sice obsazovat vzdálené lokality, ale líhnou se vyvinutější a mají tedy větší šanci na přežití..

Životní cyklus langust (infrařád Achelata): 1) dospělí jedinci žijící na útesech poblíž pobřeží; 2) po spáření a nakladení vajíček na pleopody následuje hromadná migrace samic do hluboké vody, samice nosí vajíčka na pleopodech (7-180 dní); 3) před líhnutím larev se samice vrací zpět do mělké vody; 4) planktonní larva phyllosoma (9-25 stadií, 65 dní až 2 roky); 5) částečně bentická post-larva puerulus (1 stadium, 7-56 dní); 6) juvenilní langusta (24-50 měsíců). Velikostní poměr vývojových stadií na obrázku neodpovídá skutečnosti. V závorce je u každého vývojového stadia uvedena doba, po kterou korýš v tomto stadiu setrvává
Po ukončení vývoje se bez pravé metamorfózy stává z larvy juvenilní jedinec. U suchozemských druhů v tomto stadiu obvykle opouští vodu a přesouvá se do prostředí, ve kterém žijí dospělci. U sladkovodních druhů, jejichž larvy prodělaly vývoj v moři, se juvenilní jedinci postupně posunují proti proudu řek rovněž do prostředí obývaného dospělci.
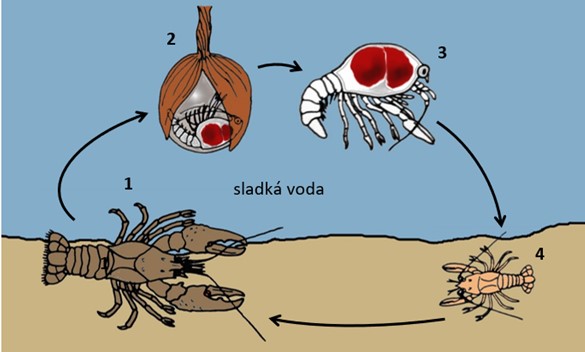
Životní cyklus raků (infrařád Astacidea): 1) gravidní samice nosí vajíčka přichycená na pleopodech; 2) většina larválního vývoje probíhá uvnitř vajíčka (7-180 dní); 3) vylíhlá mláďata jsou v bentickém stadiu, stále jsou ale přichycena k matce (3-4 stadia, 7-30 dní); 4) juvenilní rak (3 měsíce až 14 let). Velikostní poměr vývojových stadií na obrázku neodpovídá skutečnosti. V závorce je u každého vývojového stadia uvedena doba, po kterou korýš v tomto stadiu setrvává (značně se ovšem liší podle druhu a čeledi)
Vylíhlá mláďata raků odpovídají vývojem bentickým larvám ve stádiu megalopy. Přesto se v literatuře jako larvy neoznačují. Jsou stále přichycené na abdominální končetiny (pleopody) samice. U čeledí Astacidae a Cambaroididae setrvávají u matky po dvě stadia, u čeledí Parastacidae a Cambaridae jsou to stadia tři až čtyři, výjimečně ale až sedm. V prvním stadiu jsou mláďata k pleopodům matky přichycena speciálním telsonovým vláknem. Jelikož toto vlákno po několika dnech mizí, snaží se vylíhlé larvy zachytit svými kráčivými končetinami za pleopody samice či za tělo sourozenců. U čeledí Astacidae, Cambaridae a Cambaroididae k tomuto účelu slouží háčky na hrotech klepet a částečně i háčky na koncích ostatních kráčivých končetin. U druhů z čeledí Astacidae a Cambaroididae jsou tyto háčky přítomné jen u mláďat v prvním vývojovém stadiu, zatímco u severoamerických zástupců čeledi Cambaridae přetrvávají i během druhého vývojového stadia. U čeledi Parastacidae slouží k přichycení larev k matce háčky na posledních dvou párech kráčivých končetin, které přetrvávají až do ukončení druhého vývojového stadia. V druhém stadiu se mláďata začínají pohybovat. Přibližně po dvou týdnech se přeměňují na juvenilní raky, kteří postupně samici opouštějí, osamostatňují se a hledají úkryty v blízkém okolí.
Juvenilní jedinec vypadá již jako dospělý desetinohý korýš, liší se ale velikostí některých částí těla. Oproti dospělci má v poměru k tělu větší oči a menší klepeta, samice nemají ještě robustní abdomen. Celková velikost těla juvenilních korýšů závisí samozřejmě na druhu, většinou se ale pohybuje okolo 5 mm. Proces růstu a pohlavního dospívání je různě dlouhý v závislosti na druhu. Juvenilní jedinec se může změnit na dospělce již za čtyři měsíce (u raka mramorovaného, Procambarus virginalis), nebo až za devět let u samců raka tasmánského (Astacopsis gouldi), respektive až za čtrnáct let u samic tohoto druhu. Po stadiu juvenilním následuje subadultní (pohlavně nedospělý jedinec), po něm pak adultní (dospělec schopný reprodukce).
Autor textu: Doc. Ing. Jiří Patoka, Ph.D., DiS.
Autor fotografií: Doc. Ing. Jiří Patoka, Ph.D., DiS.
Publikováno:
20.3.2025 11:23
0 hlasy
Copyright 2025 © faunaportal.cz
Created by MVKV Solutions, s.r.o.